




1.TaPHL7 通过结合TaGS1;3-4D启动子抑制其转录活性
研究者以TaGS1;3启动子为诱饵,通过酵母单杂交(Y1H)对小麦籽粒 cDNA 文库进行筛选,鉴定MYB家族候选转录因子TaPHL7。通过 Y1H、蛋白凝胶阻滞(EMSA)与染色质免疫沉淀和定量PCR (ChIP-qPCR)证明它能够识别P1BS基序与TaGS1;3-4D启动子结合,通过双荧光素酶报告基因检测试验(Dual-Luciferase Assay)进一步证明 TaPHL7 负向调控TaGS1;3-4D启动子的表达。表明 TaPHL7 通过识别 P1BS 基序结合TaGS1;3-4D启动子抑制其转录活性(图1)。
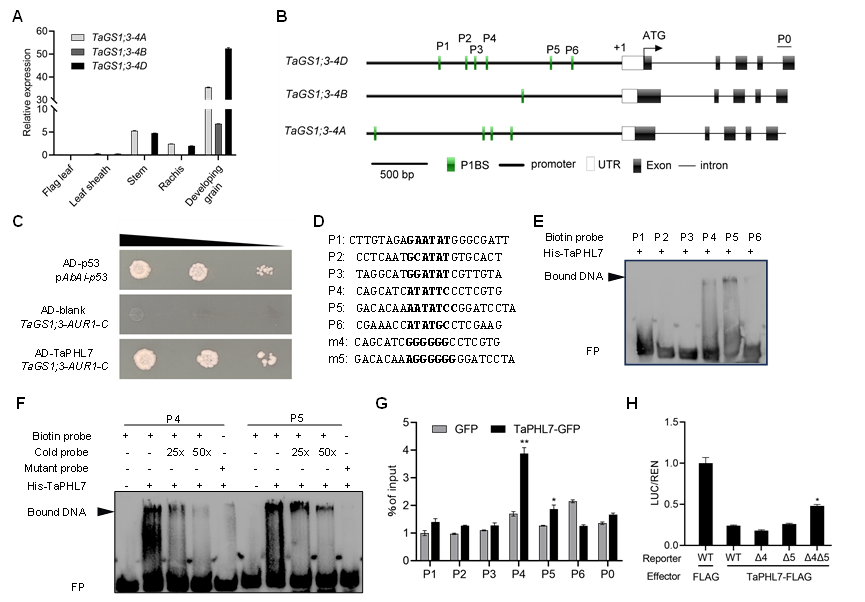
图1:TaPHL7 通过结合TaGS1;3-4D启动子抑制其转录活性
2.TaPHL7调控植物生长与发育
利用遗传转化技术获取TaPHL7过表达和 CRISPR-Cas9 介导的突变 T3 代纯合株系后,将遗传材料在北京、河南两地种植,对其农艺性状调查显示,在幼苗营养生长期taphl7突变体相比较野生型出现窄叶表型;在抽穗期,taphl7 突变体相比较野生型株高增加,分蘖增多,抽穗日期延后 4-5 d;但呈现早熟表型,单株产量增加;而TaPHL7过表达植株中与野生型无显著差异。表明 TaPHL7 在调节植物生长发育中发挥重要作用(图2)。
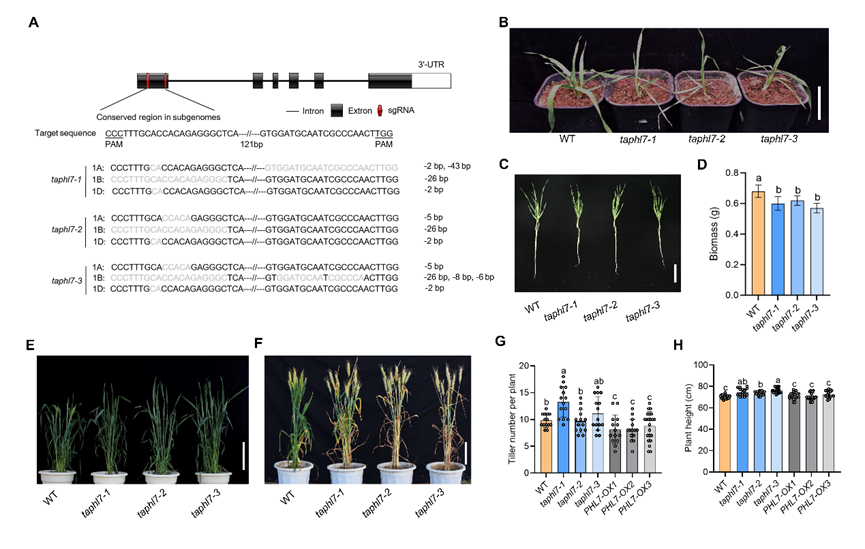
图2:TaPHL7调控植物生长和发育
3.TaPHL7 正向调控 Pi 信号传导和 Pi 吸收
研究者为了探究TaPHL7在磷信号转导中的作用,水培条件下用浓度分别为 0.20 mM KH2PO4(正常磷,NP)和 0.01 mM KH2PO4(低磷,LP)检测了小麦野生型ZM7698 及taphl7突变体植株中的磷反应。在正常磷或低磷条件下,TaPHL7 功能缺失抑制侧根发育,降低磷吸收与转运能力。此外,qRT-PCR 结果表明,TaPHL7功能缺失导致磷饥饿响应基因PSR(TaPHT1;9、TaSPX3)的表达在低磷条件下显著下调,表明TaPHL7 功能缺失后导致磷信号转导通路受损。ChIP-qPCR 及双荧光素酶试验进一步证实,TaPHL7 通过结合磷转运蛋白基因TaPHTs(TaPHT1;9、TaPHT1;3)启动子,正向调节 TaPHTs的表达,促进磷的获取(图3)。
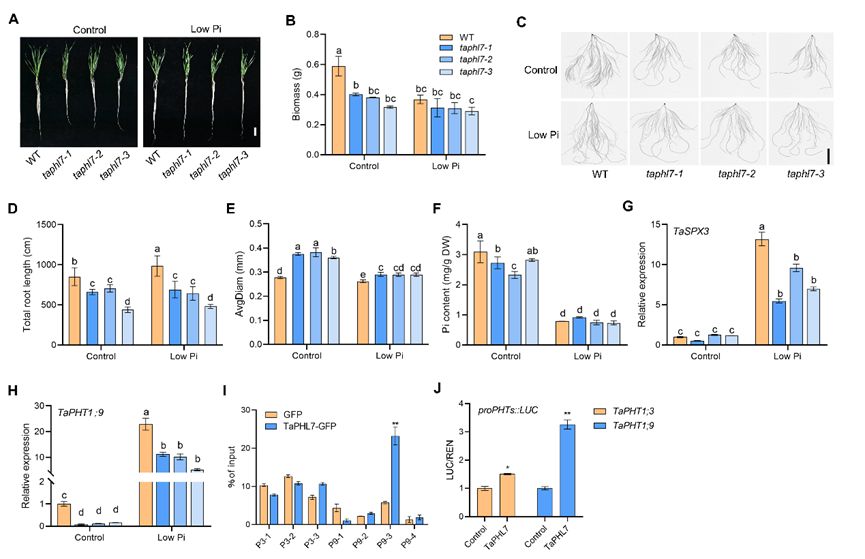
图3:TaPHL7 正向调控 Pi 信号传导和 Pi 吸收
4.TaPHL7 负调控氮代谢
TaPHL7功能缺失在开花期旗叶中表现为总氮含量增加,灌浆期籽粒中 GS 酶活性增加,氮同化及再动员能力增强;而过表达TaPHL7的植株则与此相反。稳定同位素试验表明:TaPHL7功能缺失促进了幼苗期硝酸盐的吸收、转运能力。表明 TaPHL7 在营养生长及生殖生长阶段均负向调控氮的吸收、转运及再分配(图4)。
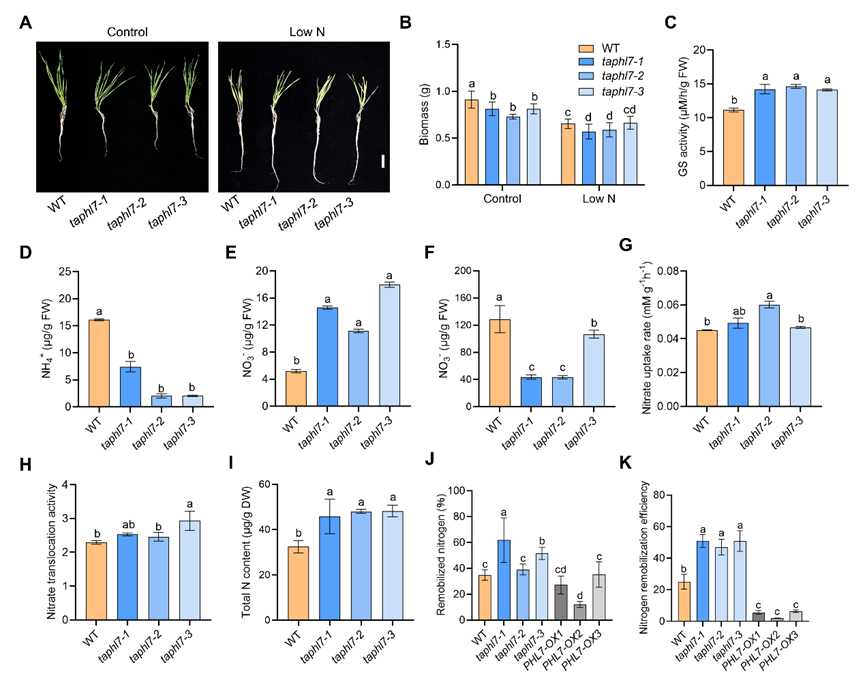
图4:TaPHL7 负调控氮代谢
5.TaPHL7调控的氮磷利用调节籽粒产量
接下来研究者探讨了TaPHL7是否参与调控氮-磷信号通路以影响植物生长发育。TaPHL7通过不同的机制调控氮和磷的信号传导:硝酸盐可能通过解除TaPHL7对TaNRT2.1和TaNRT2.2的转录抑制来发挥作用,同时直接促进TaPHT1;3和TaPHT1;9的表达。由于TaPHL7可能调控氮和磷的利用,研究者进行为期两年的田间试验中探讨了其在籽粒产量方面的潜力。TaPHL7功能缺失提高了单株产量、穗粒数和千粒重。这些结果表明,在自然栽培条件下,TaPHL7功能缺失突变可促进早熟并提高籽粒产量(图5)。
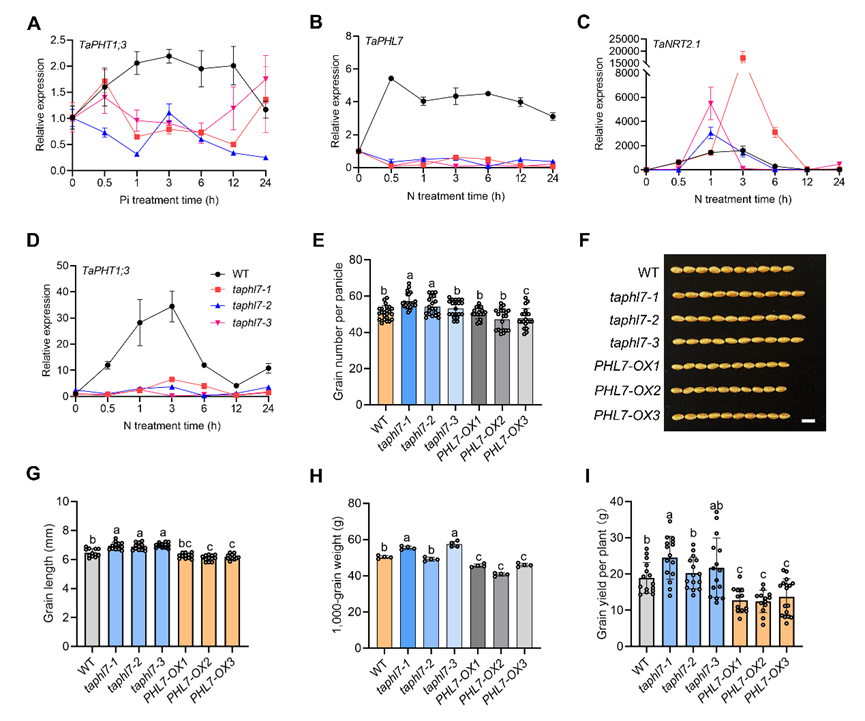
图5:TaPHL7调控的氮磷利用调节籽粒产量
我院马新明教授、中国科学院遗传与发育生物学研究所左建儒研究员为本论文的共同通讯作者,我院博士生王华丽为论文第一作者,张志勇副教授为论文的共同第一作者。河南农业大学生命科学学院王小纯教授,农学院熊淑萍教授,中国科学院遗传与发育生物学研究所鲁非研究员,中国农业大学农学院倪中福教授,姚颖垠教授对本研究工作给予了指导和帮助。本研究得到国家自然科学基金委员会(32071956、32330010)、中华人民共和国科学技术部(2021YFD1700900、2022YFD1201700)和植物基因组学国家重点实验室(SKLPG2023-22)的资助。