


学院新闻
植物长期遭受干旱、盐碱、低温、病虫害等多重胁迫,产量与稳定性受限。植物依靠激素信号、MAPK 级联反应、转录重编程等协同应对逆境。MYB 转录因子是植物最大转录因子家族之一,功能多样,但其如何协同调控多种激素信号、协调逆境响应与代谢调控的系统机制尚不清晰。
2026年6月1日,我院小麦分子育种创新团队在植物学国际著名期刊Plant Biotechnology Journal上发表了题为“MYB Factors: Hubs of Plant Stress and Hormone Crosstalk”的长文综述。该文章以 MYB 转录因子为核心,系统阐明其作为植物逆境信号与多种激素通路的 “调控枢纽”,整合植物生物非生物胁迫、激素信号、次生代谢及生长发育的分子机制,为培育抗逆作物新品种提供重要理论依据。
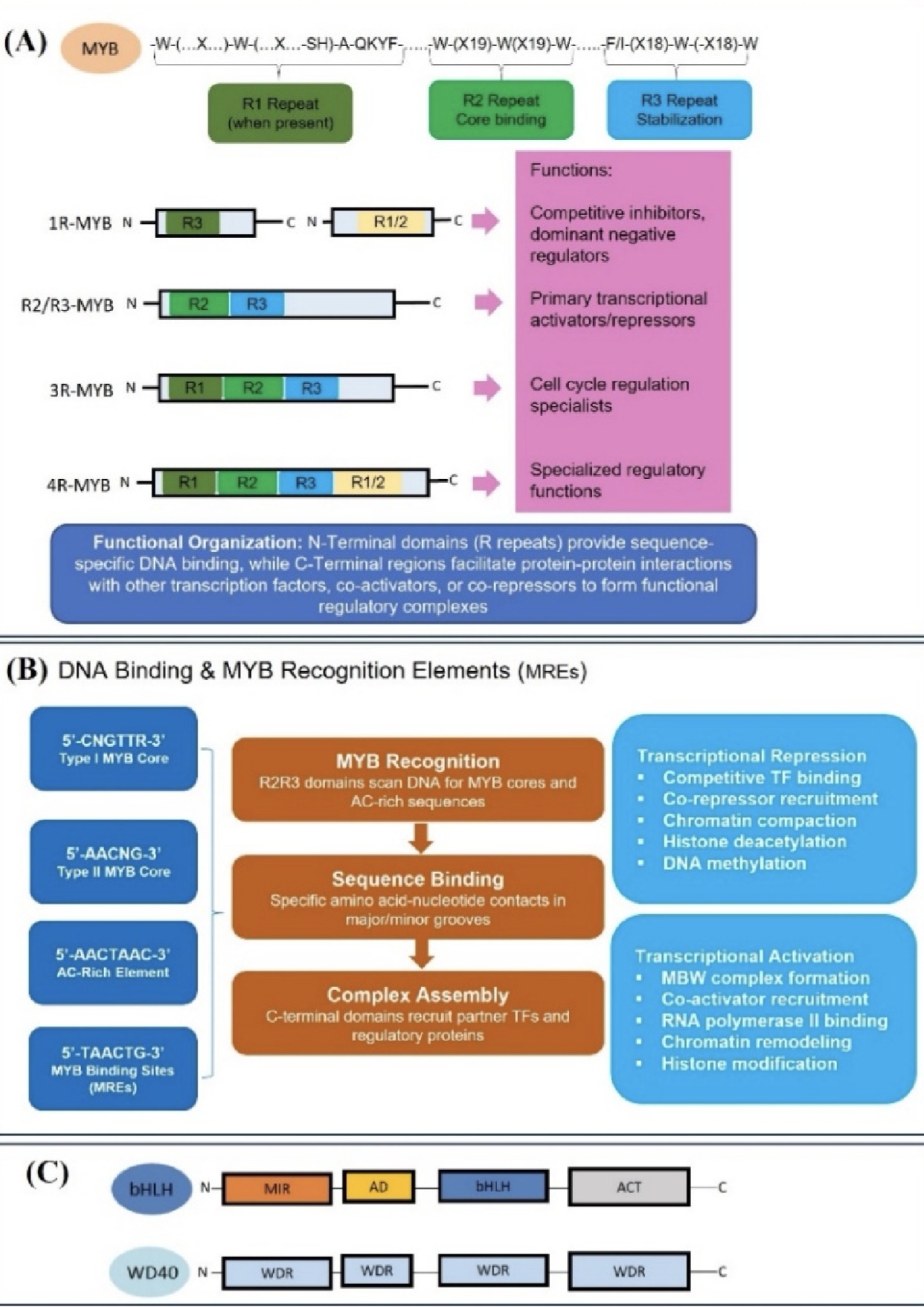
 植物MYB转录因子分为1R-MYB、R2R3-MYB、3R-MYB、4R-MYB四类,该家族蛋白结构特征鲜明(图1),N端相对保守,通过DNA序列与蛋白结合;C端序列变异丰富,决定家族成员的功能分化。在结构功能层面,R2R3-MYB可识别MRE与富AC顺式元件,通过蛋白互作整合多条信号通路。抑制型MYB的C端含多种保守基序,且功能特异。MYB可靶向结合靶基因启动子,调控基因转录。MYB可与bHLH、WD40形成MBW三元复合物,调控植物代谢、抗逆与生长发育。该复合物结构保守,可通过磷酸化修饰、结构转换、可逆组装实现动态调控,适配植物生理与环境变化。
植物MYB转录因子分为1R-MYB、R2R3-MYB、3R-MYB、4R-MYB四类,该家族蛋白结构特征鲜明(图1),N端相对保守,通过DNA序列与蛋白结合;C端序列变异丰富,决定家族成员的功能分化。在结构功能层面,R2R3-MYB可识别MRE与富AC顺式元件,通过蛋白互作整合多条信号通路。抑制型MYB的C端含多种保守基序,且功能特异。MYB可靶向结合靶基因启动子,调控基因转录。MYB可与bHLH、WD40形成MBW三元复合物,调控植物代谢、抗逆与生长发育。该复合物结构保守,可通过磷酸化修饰、结构转换、可逆组装实现动态调控,适配植物生理与环境变化。
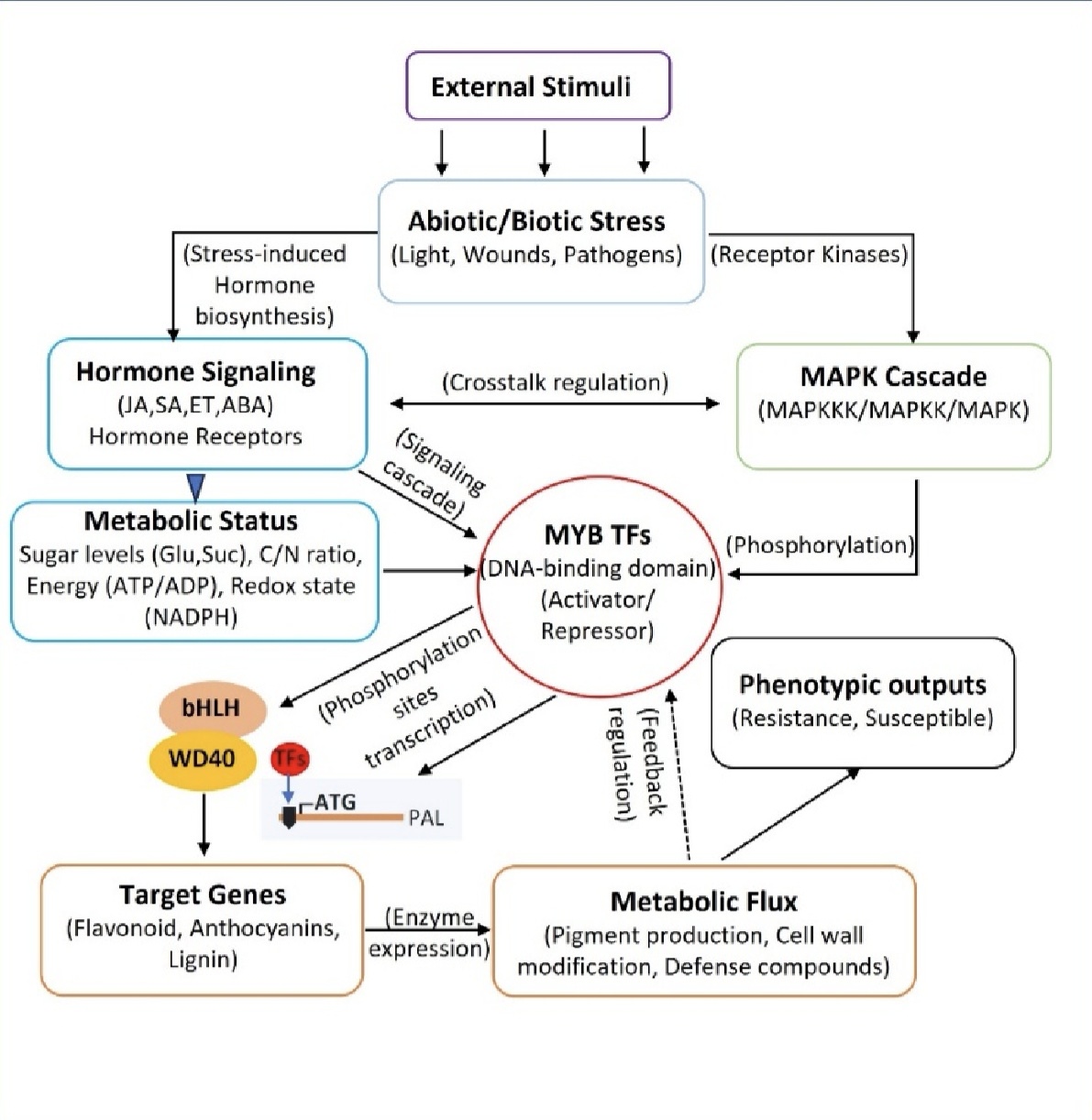
总之,本文系统构建了从胁迫信号、激素/MAPK通路、MYB,到MBW复合物,最终到代谢/防御响应的调控模型,证明MYB转录因子是植物应对多重逆境的核心信号枢纽,不仅协调激素交叉对话与次生代谢,更平衡生长与防御,为培育气候智慧型作物提供了关键理论支撑与技术方向。
我院博士留学生Aye Aye Htun为该论文第一作者,我院陈锋教授为通讯作者。本研究得到国家重点研发计划(2025YFE0209700)和国家自然科学基金项目(W2412001)资助。
文章链接:https://doi.org/10.1111/pbi.70694
2026年6月1日,我院小麦分子育种创新团队在植物学国际著名期刊Plant Biotechnology Journal上发表了题为“MYB Factors: Hubs of Plant Stress and Hormone Crosstalk”的长文综述。该文章以 MYB 转录因子为核心,系统阐明其作为植物逆境信号与多种激素通路的 “调控枢纽”,整合植物生物非生物胁迫、激素信号、次生代谢及生长发育的分子机制,为培育抗逆作物新品种提供重要理论依据。
图1. MYB 转录因子的结构与功能基础
研究发现,MYB转录因子广泛参与植物生物胁迫与高温胁迫应答。小麦遭遇病原菌侵染或高温胁迫时,TaMYB73特异性诱导表达,通过上调TaPR1、TaPR2等水杨酸通路关键防御基因,激活植株水杨酸抗性通路。其中MYB30可介导不依赖NPR1的水杨酸防御反应,是植物抗逆体系中的独特功能因子。
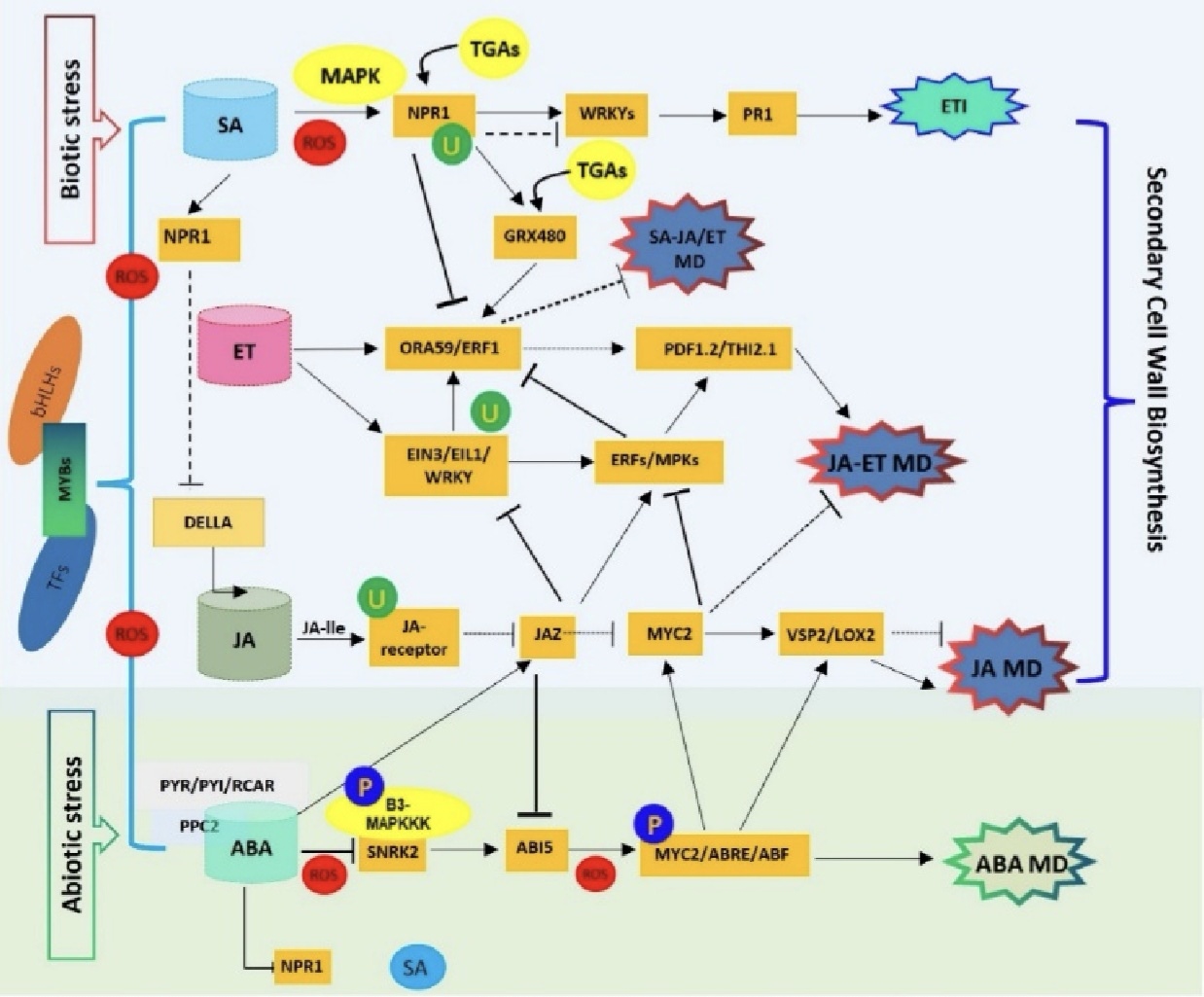
图2. MYB转录因子是SA、JA、ET、ABA四大激素通路的信号核心枢纽
研究证实,各类植物激素主要通过隔离抑制因子间接调控MBW转录复合物,其中乙烯的调控效应存在显著的条件依赖性与物种差异。拟南芥中,乙烯信号蛋白EIN3可阻断TTG1与GL3的互作及GL2基因表达,抑制MBW复合物组装,并通过RSL4基因调控表皮毛与根毛发育,重塑植株表皮形态结构。梨中,乙烯通过PpERF9招募共抑制因子PpTPL1,促进组蛋白去乙酰化,从而抑制花青素合成关键因子PpMYB114及其上游调控基因PpRAP2.4的表达,进而负向调控花青素积累。研究表明,茉莉酸依托MBW复合物的动态组装调控植物次生代谢,且该调控通路具有明显物种特异性。JAZ家族蛋白可竞争性结合GL3、EGL3、TT8等bHLH核心蛋白,阻碍MBW复合物形成,抑制类黄酮合成。苹果存在独特的JAZ1-TRB1-MYB9花青素调控模块,区别于其他物种,直观体现MYB-JAZ通路的物种分化特征。苹果中MdERF1B可激活MYC2与R2R3-MYB家族基因,低茉莉酸水平下,MdJAZ5/10会抑制其活性;而茉莉酸可诱导JAZ蛋白泛素化降解,解除抑制作用,促进MBW复合物组装与花青素合成。杨树遭受损伤胁迫时,JAZ1蛋白降解可激活MYB118-TT8复合体,诱导花青素合成基因表达,启动胁迫防御。以外,赤霉素与茉莉酸通路可协同调控植物花青素合成。赤霉素信号通路中的DELLA蛋白能够结合并隔离MYBL2、JAZ等MBW复合物拮抗因子,释放MYB、bHLH核心功能亚基,促进花青素合成。MYB30介导的激素调控通路在植物中功能保守。番茄中SlMIEL1既可介导MYB30同源蛋白泛素化降解,也可降解茉莉酸合成负调控因子SlNAC72,解除其对SlLOX10、SlAOS1、SlAOC等茉莉酸合成基因的抑制,提升内源茉莉酸含量、清除活性氧,增强植株高温耐受性。逆境与脱落酸处理可提升MYB30、ABI5蛋白稳定性,降低MIEL1蛋白积累,进一步强化植物胁迫应答能力。综上所述(图2),能量胁迫可通过蛋白翻译后修饰重塑MYB调控网络,改变植株代谢模式。能量胁迫激酶SnRK1可磷酸化修饰MYB75、TT8、TTG1等MBW核心组分,引发复合物解离、MYB75降解,抑制DFR、UFGT等花青素合成关键基因表达,促使植株由生长合成代谢转向逆境适应代谢,从而提升植株胁迫存活能力。
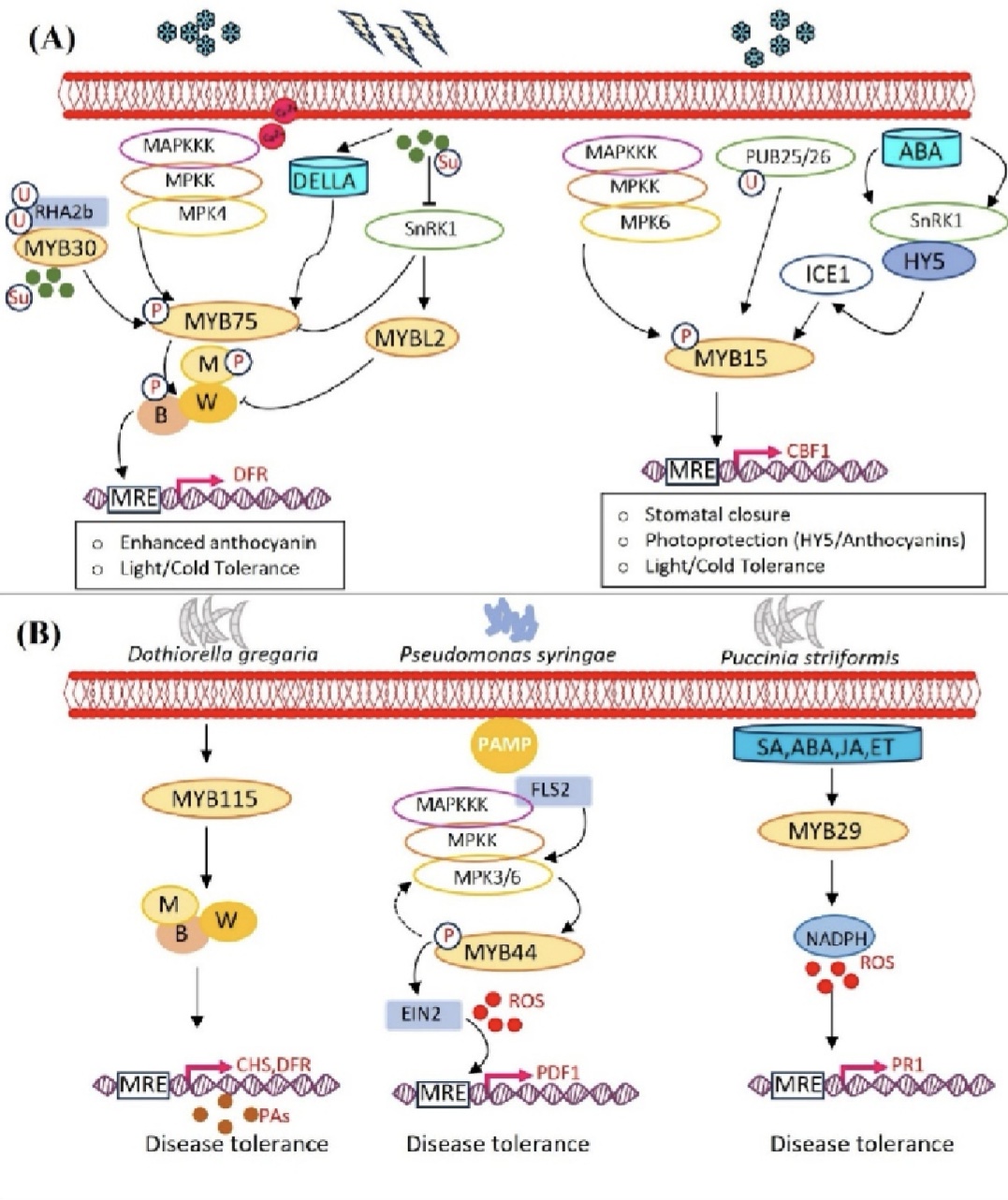
图3. MYB是多重逆境信号整合载体
拟南芥花青素合成受多信号通路与蛋白互作网络精细调控,MYB30可与MYB75互作并阻碍其组装MBW复合物,从而抑制花青素生成,而蔗糖可激活MYB75介导的RHA2b依赖型MYB30泛素化降解,解除抑制并促进花青素积累,同时SnRK1可介导MYB75降解以阻断下游结构基因转录,赤霉素DELLA蛋白则通过隔离MYBL2与JAZ抑制因子正向促进花青素合成。MYB家族广泛参与冷、盐等非生物胁迫调控,拟南芥PUB25/26可通过泛素化修饰动态调控ICE1、MYB15稳定性,且MPK6介导的磷酸化可弱化MYB15对CBF3的抑制作用,精细调控冷信号通路。在免疫防御层面,MYB通过调控苯丙烷代谢维持植物抗病平衡,MYB115依托MBW复合体正向调控原花青素合成,显著增强植株真菌抗性。此外,在病原相关分子模式(PAMP)触发的植物免疫反应中,MPK3 与 MPK6 激活后会启动下游信号级联通路,促使拟南芥MYB44 蛋白发生瞬时磷酸化;磷酸化修饰后的 MYB44 可激活下游防御基因,并诱导乙烯信号关键基因EIN2的表达。小麦TaMYB29可整合水杨酸、茉莉酸、脱落酸、乙烯多重激素信号,通过促进过氧化氢积累、激活病程相关基因表达,强化植株过敏免疫反应与条锈病抗性,是激素介导植物免疫的核心正向调控因子。
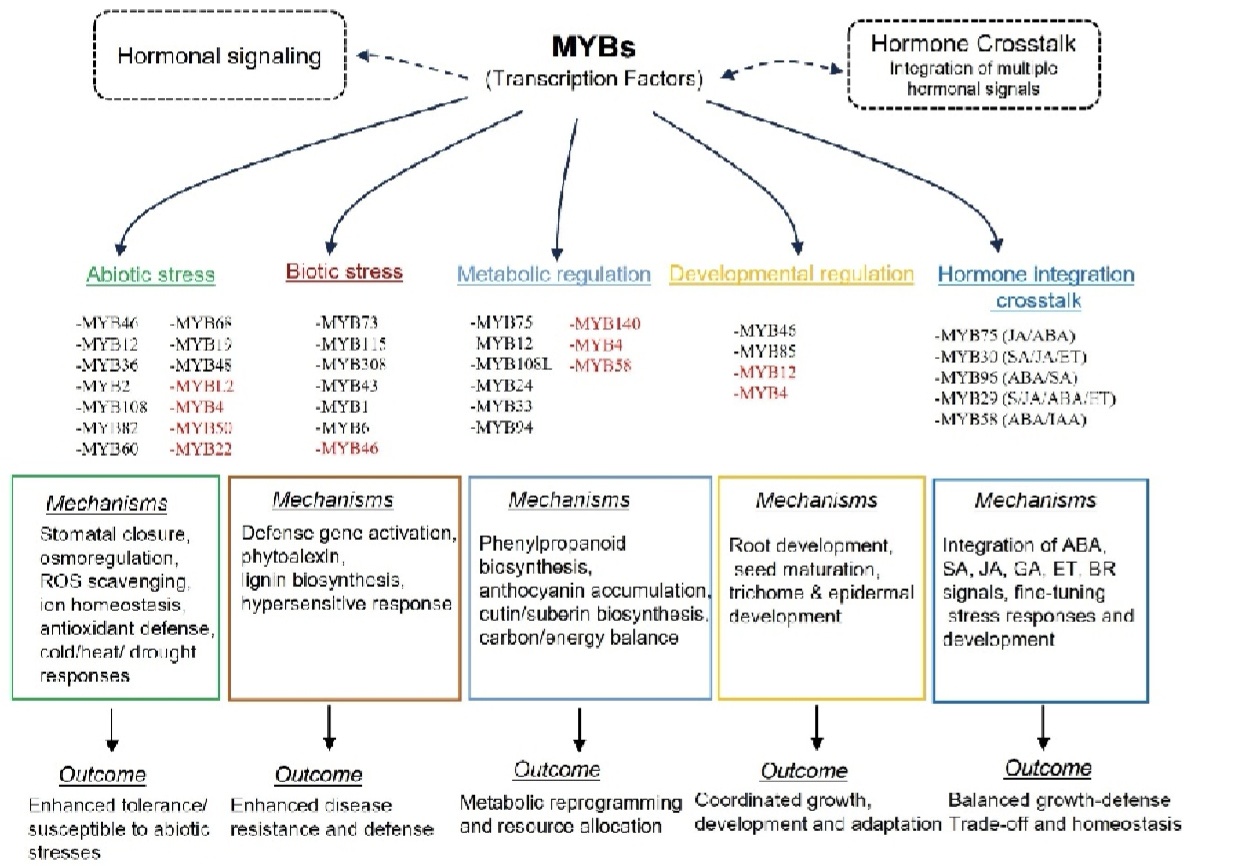
图4. MYB 转录因子协调植物中的非生物胁迫、生物胁迫、代谢及发育进程。
MYB4转录因子存在显著物种功能分化。苹果中MbMYB4可增强植株抗旱能力,而含EAR基序的VcMYB4a则抑制胁迫基因表达,加剧干旱、盐胁迫敏感性。拟南芥AtMYB4、AtMYB7、AtMYB32可竞争结合bHLH蛋白,抑制苯丙烷代谢。不同物种MYB4功能差异明显:MbMYB4提升耐旱性,VcMYB4a增强胁迫敏感性,PvMYB4调控木质素代谢,小麦TaMYB4负调控耐冷性。AtMYB4依赖GIR蛋白入核,可独立调控生长与代谢,动态平衡植株生长与逆境适应。同时,MYB转录因子可通过调控细胞壁重塑参与植物逆境适应。柚CgMYB58可整合脱落酸与生长素的拮抗信号,调控木质素合成相关基因表达,调节细胞壁木质素积累,介导植株逆境结构适应性调控。此外,MYB46、MYB85保守调控植物细胞壁代谢与免疫防御。MYB85协同MYB20/42/43激活木质素与苯丙烷合成,其功能缺失会抑制植株生长、削弱次生细胞壁发育。烟草、竹子MYB85可促进木质素沉积以加固细胞壁;拟南芥AtMYB46靶向调控Ep5C,敲低其表达可重塑细胞壁、增强灰霉菌抗性,关联细胞壁重塑与免疫应答。MYB96为多重胁迫核心调控因子,小麦TaMYB96-2D通过调控蜡质合成减少干旱失水。拟南芥AtMYB96可通过蜡质积累与根系改良提升抗旱性,结合HDA15增强ABA信号,调控碳脂代谢与逆境种子萌发,同时促进水杨酸合成,同步调控生物与非生物胁迫抗性。综上,MYB转录因子功能多样,广泛参与植物胁迫应答、次生代谢、生长发育与激素信号整合如(图4)所示,通过精密调控网络平衡植株生长、代谢稳态与逆境防御,是调控植物环境适应性的核心转录因子家族。
图5. MYB协调介导应激反应与代谢模型
植物免疫应答中,PAMP信号可激活MPK3/MPK6,介导MYB44瞬时磷酸化,从而激活下游防御基因与EIN2表达;同时,渗透胁迫下MPK3下游的VIP1可特异性调控MYB44活性。MPK6兼具生长与逆境调控双重功能,其在木质部发育中持续激活,通过介导MYB46磷酸化降解,精准调控次生细胞壁合成。此外,光照、低温及氧化胁迫可激活MPK4,对MYB75多个位点进行磷酸化;其中Thr126、Thr131位点磷酸化可阻断COP1结合,抑制MYB75降解,进而稳定MYB75蛋白,促进花青素积累,正向提升植株低温与氧化胁迫抗性。综上,MAPK通过特异性磷酸化动态调控MYB蛋白稳定性与活性,广泛参与植物免疫、细胞壁发育与非生物胁迫应答,是平衡植物生长与逆境适应的核心调控模块。总之,本文系统构建了从胁迫信号、激素/MAPK通路、MYB,到MBW复合物,最终到代谢/防御响应的调控模型,证明MYB转录因子是植物应对多重逆境的核心信号枢纽,不仅协调激素交叉对话与次生代谢,更平衡生长与防御,为培育气候智慧型作物提供了关键理论支撑与技术方向。
我院博士留学生Aye Aye Htun为该论文第一作者,我院陈锋教授为通讯作者。本研究得到国家重点研发计划(2025YFE0209700)和国家自然科学基金项目(W2412001)资助。
文章链接:https://doi.org/10.1111/pbi.70694