


学院新闻
小麦是全球的主食,也是种植和消费最广泛的作物之一。鉴于全球人口不断增长和对小麦的需求不断增加,提高小麦产量至关重要。籽粒大小通过影响粒重来影响产量。然而,由于小麦基因组庞大而复杂,利用基于图谱的克隆策略直接挖掘产量相关基因具有挑战性。目前,通过不同组学、EMS突变体等方法克隆出了一些产量相关基因。但受体激酶(LecRLKs)作为植物特异性的RLK亚家族,参与植物发育、种子萌发、激素信号传导等,目前在调控小麦籽粒大小中的作用还尚未可知。
近日,我院陈锋教授团队在Science Bulletin(即时IF=20.6)发表了题为“A lectin receptor-like kinase TaSRK that is deacetylated by TaHDA9 regulates wheat grain length by mediating the Photosystem II protein TaPsbO”的研究论文。该研究在小麦中克隆到一个负向调控小麦籽粒长度的关键基因TaSRK, 并解析了TaHDA9-TaSRK模块通过介导光系统II中的TaPsbO调控小麦籽粒长度的分子机制。


TaHDA9-TaSRK-TaPsbO模块通过介导光合调控小麦籽粒长度
近日,我院陈锋教授团队在Science Bulletin(即时IF=20.6)发表了题为“A lectin receptor-like kinase TaSRK that is deacetylated by TaHDA9 regulates wheat grain length by mediating the Photosystem II protein TaPsbO”的研究论文。该研究在小麦中克隆到一个负向调控小麦籽粒长度的关键基因TaSRK, 并解析了TaHDA9-TaSRK模块通过介导光系统II中的TaPsbO调控小麦籽粒长度的分子机制。
河南农业大学陈锋教授团队发表研究论文
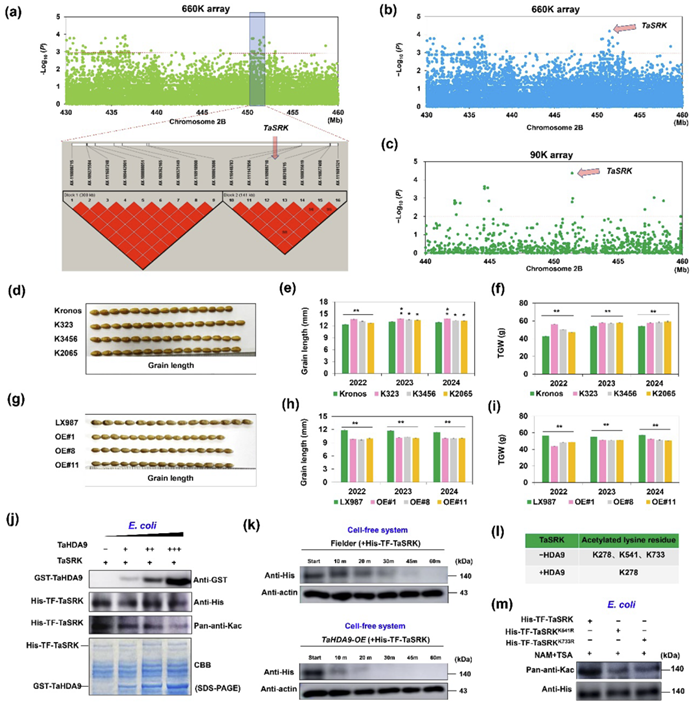
为了挖掘小麦籽粒大小调控基因,本研究对349份小麦材料进行了多年多点的籽粒大小和千粒重调查,通过GWAS分析确定一个多环境显著的Block、该区间包含了11个候选基因,并进一步结合EMS突变体库、转录表达和小麦Pan800K的基因测序信息等锁定TaSRK为关键候选基因(图1b-c),该基因存在4种单倍型,其中TaSRK-Hap1的粒长和千粒重显著较高。连续3年田间数据显示,TaSRK过表达转基因小麦粒长变短、千粒重显著下降,而突变体则显示粒长变长、千粒重显著上升(图1d-i)。以TaSRK蛋白为诱饵,通过酵母双杂交筛选小麦cDNA文库,确定了一个组蛋白去乙酰化酶TaHDA9。实验证实TaSRK和TaHDA9存在蛋白互作。体外和半体外去乙酰化实验均证实TaHDA9直接去乙酰化TaSRK的2个赖氨酸位点K541和K733,并由此降低其蛋白积累(图1j-m)。2022-2024连续3年农艺性状调查结果表明,TaHDA9过表达转基因小麦籽粒变长、千粒重显著增加,而突变体籽粒变短、千粒重显著下降。
TaSRK的克隆及功能鉴定
为了阐明TaSRK调控小麦粒长的机制,我们还重点研究了另一个筛库蛋白TaPsbO。实验证实TaSRK在叶绿体中与TaPsbO相互作用,表明TaSRK可能通过与叶绿体中的TaPsbO相互作用来发挥其功能。体内外磷酸化结果表明,TaSRK可以直接磷酸化TaPsbO(TaPsbOS222)的7个位点(包括关键位点S222)并降低其蛋白丰度(图2a-c)。单倍型分析结果显示,TaPsbO在自然界中存在一个稀有单倍型 TaPsbOG222。蛋白互作结果表明,TaSRK与TaPsbOG222互作更强。体内外磷酸化实验证实TaSRK对TaPsbOS222和TaPsbOG222存在差异磷酸化调控,且TaPsbOG222单倍型蛋白丰度更高。随后通过体内外的三者实验对TaHDA9-TaSRK-TaPsbO模块调控小麦粒长的机制进行了解析,证实TaHDA9通过去乙酰化调控TaSRK进而促进TaPsbO的蛋白积累(图2d-f)。通过小麦过表达植株及原生质体水平实验证实,TaPsbO的表达分别受TaHDA9和TaSRK调控(图2g-j)。连续3年田间农艺性状调查数据显示,TaPsbO-RNAi和EMS突变体小麦籽粒变短、千粒重显著降低。进一步测定上述3个基因过表达和突变体材料的光合相关参数(Fv/Fm, ETRII, chlorophyll含量和CO2 同化率),结果显示,这些材料的光合参数均出现显著变化,表明这3个基因与光合调控密切相关。连续3年对TaSRK过表达和突变体进行了单株产量,小区产量的测定,结果显示,与对照相比,突变体产量显著增加,过表达产量显著下降。综上所述,本研究挖掘到一个调控小麦粒长的新基因TaSRK,并阐明了TaHDA9-TaSRK模块通过介导光系统II中的TaPsbO来增强小麦粒长的级联调控机制。
TaHDA9-TaSRK-TaPsbO模块通过介导光合调控小麦籽粒长度